

Evolving the Irreducible
Behe's Mousetrap Problem
Sean D. Pitman M.D.
© October 2008
Michael Behe, a professor of biochemistry at Lehigh University, put himself out on a line in his book, Darwin’s Black Box, claiming that there are systems of function in the biological world that are "irreducibly complex." As an example for the layperson who might not understand biochemical models, Behe used a common mouse trap as an example of irreducible complexity. Such a trap requires a minimum number of parts to work. These include a platform, spring, hammer, hold-down bar, catch, and staples. If any one of these parts is taken way, the mousetrap will not work at all, much less a little bit.2 Obviously, this is an irreducibly complex system - right? How could such a system evolve gradually if all parts had to arrive on the scene suddenly? Ah ha! Evolution is in a real fix now! ( Back to Top )
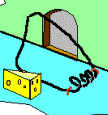
Not so fast. Professor John H. McDonald from the University of Delaware challenged the notion that Behe’s mousetrap is irreducibly complex. He proposed a stepwise evolution of Behe’s mousetrap starting with a wire loop held open under tension. From this simple and fairly ineffective wire loop, McDonald demonstrated a fairly convincing series of small evolutionary changes that ended up turning into Behe’s "irreducible" mousetrap. McDonald explains each successive step in the evolution of the mousetrap using a single mutational event to produce a small and yet functional change to the previous mousetrap. Consider McDonald’s illustrations below.9
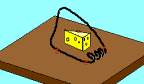
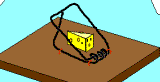
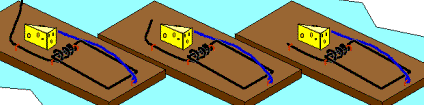
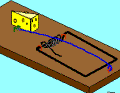
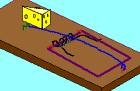

A wire is held open under tension. A coil is added to provide greater force. Bait is added to better tempt the mouse. Clips are added to secure the trap in one place. The bait is moved to a more strategic location. The hammer wire is lengthened and secured to provide more surface area and stability. A hold-down bar is added to provide more force to the hammer bar. The hold bar is moved to a clip to provide more trap durability. Extra wire that is no longer serving a function is shortened. The hold-down bar gets lodged under a clip instead of the hammer bar to improve its release potential. Adding a swiveling catch to the clip improves the release potential of the hold-down bar even more. And presto, the irreducibly complex trap has evolved from simple to complex.
Pretty convincing isn’t it? Has McDonald saved the theory of evolution from the problem of irreducible complexity? It sure seems like he has indeed. But, is the problem of irreducibly complexity really this easy to overcome? The very fact that McDonald and so many other prominent evolutionists vehemently argue against the very existence of irreducible complex systems seems to indicate that this concept is in fact a significant challenge to the theory of evolution - if it were true. If theory of evolution is true, then "irreducible" systems do not exist in the natural world in such a way that the steps required to form such systems cross anything more than very small gaps of neutral/non-beneficial function. Therefore, those who accept the truth of evolution, cannot accept the idea that any biosystem structure with a beneficial function, however complex, is very far removed from the next closest functional system or subsystem within the potential of "sequence space" that might be beneficial if it were ever found by random mutations of any kind. ( Back to Top )
Evolutionists do in fact put great effort into proposing all sort of potential evolutionary pathways to explain the existence of complex multi-part biological systems that are vastly more involved than Behe's mousetrap illustration. For example, consider Dawkins’s computer model for evolutionary mechanisms producing the phrase, “Methinks it is like a weasel” starting with a random sequence over a few "generations". Dawkins showed that his computer could evolve this phrase in a very short time using the forces of random mutation and selection. However, Dawkins had a problem that McDonald did not have.
Dawkins evolved a meaningful phrase by starting with a random meaningless sequence of characters. At least McDonald starting with a mousetrap with at least some beneficial function. Dawkins programmed his computer to select based on phrase comparison to the line from Hamlet, "Methinks it is like a weasel." Every mutation that produced a sequence that was closer to this phrase in spelling was chosen by the computer as the most reproductively "fit." 1 In just a few dozen generations the "Methinks . . . " target phrase was realized by the "population" of evolving sequences.
Now, does this model of how evolution works actually reflective of how evolutionary mechanisms actually work in real life? - with real biosystems? Actually, there seem to be several examples of this sort of evolution happening in real biosystems. For example, the immune system demonstrates this sort of evolution. The antibodies that are produced by B-cells are modified over a few generations of B-cells via random mutation and function-based selection. Those B-cells that produce antibodies that match a particular foreign antigen sequence, as part of an invading organism, are given a reproductive advantage over their peers. In each generation, random mutations to the produced antibody sequence end up producing a range of possibilities, with some antibodies within that range having a greater match to the foreign antigen sequence than did the parent generation - just like what happened in Dawkins's model. Very quickly, a high level of antigen specificity is produced with the use of random mutation and function-based selection.
Another example comes in the form of experiments done by Hayashi et al. What Hayashi was wondering is if a virus that is usually able to infect a particular type of bacteria would still be able to be infective if its binding sequence were replaced by a random protein sequence. Sure enough, the virus was able to bind to the bacterium using a completely random sequence - although much much less effectively. However, like McDonald's illustration of an evolving mousetrap described above, Hayashi's viruses quickly evolved back to their original wild-type level of infectivity with the use of random mutations and function-based selection. Each generation of viruses experiences random mutations to their bacterial pilus binding sequences. Those mutations that resulting in better binding resulted in improved reproductive potential for those viruses that happened to realized these fortunate mutations. In just a few generations the original wild-type level of infectivity was regained.10 Again, this is just like what Richard Dawkins demonstrated with his computer model. ( Back to Top )
The problem, of course, is that nature cannot always produce novel beneficial functions by comparing genetic sequences to a pre-established sequence. For example, how would a bacterium evolve a function like a single protein enzyme? - like a lactase enzyme? A lactase enzyme is a protein that is able to dramatically improve the break down of the sugar lactose into glucose and galactose - each of which is then able to enter the glycolytic pathway to be used for energy. But, how would a bacterium evolve the lactose function? It is a simple matter of getting a random protein sequence to bind to a lactose molecule more and more firmly? Not really. Simple binding is not enough. The lactase enzyme has to bind in a fairly specific way to the lactose sugar in order to hydrolyze it to a useful degree. Until this threshold is reached, there simply is no improved reproductive advantage gained by the bacterium. It seems that this threshold requirement is about 380 fairly specified residues at minimum. A useful lactase simply cannot be made with significantly lower minimum size and specificity requirements.
These minimum requirements create a kind of threshold beyond which the lactase function simply cannot be built up gradually where very small one or two residues changes at a time result in a useful change in the degree of the lactase function. Therefore, such functions cannot be evolved with Dawkins's or McDonald's proposed model. There simply is no template or gradual step by step pathway from just any starting point to the minimum threshold requirement. Only after this threshold has been reached can evolution take over and make further refinements - but not until.
Now, there are in fact examples of computer evolution that attempt to address the above problem, but how would someone like McDonald deal with this problem when it comes to evolving real biosystems? ( Back to Top )
I am not exactly sure what McDonald would propose, but maybe he would start with a functional phrase of some sort and then evolve the phrase, “Methinks it is like a weasel,” in a stepwise fashion with each change maintaining function along the way. Now, this sort of thing is rather easy if one is using small character sequences - like those with only, say, three characters - as in cat to hat to bat to bad to bid to did to dig to dog. See, that wasn't too hard. Each and every mutation in this pathway produced a new sequence with a new meaning (don't worry about the "beneficial" nature of the meaning just yet). But, is it as easy to do with longer and longer sequences?
Lets try it and see. Lets start with the phrase, “Give me freedom or give me death.” In Dawkins’s illustration, a mutational event changed only one character at a time. Doing it that way would be very challenging indeed. Just try it and you will soon find that it is extremely difficult to change just one letter at a time and evolve much of anything while maintaining the meaning or "function" of the complete phrase the entire time (much less a "beneficial" function).
The reason for this increase in difficulty has to do with the nature of all symbolically represented information systems - like English, computer codes, and, yes, even genetically based biosystems. Consider that the ratio of defined vs. non-defined 3-character sequences in the English language system is about 1 in 18 (out of the potential of 17,576 three-letter sequences). Compare this ratio to the ratio for 7-character sequences of about 1 in 250,000 (out of over 8 billion). This exponential decrease in ratio makes it more and more difficult to find single character differences between what is and what might be. The same thing is true when it comes to many types of meaningful/beneficial biosystem functions. Those that require greater minimum threshold sequence size and specificity are much harder to find than those that have a lower minimum threshold requirement.
For
the
sake of illustration, lets say that we can change whole words instead of just single
characters or letters.
This is a bit easier (especially if we don't have to worry about the
function of our evolving phrases being "beneficial" during each step).
Our first evolutionary step could read, “Give me freedom or give me
death weasels.”
Then, “Give me freedom or give me weasels.”
Then, “Give me freedom or give me likeable weasels.”
Then, “Give me freedom or give like weasels.”
Then, “Give me freedom or give like the weasels.”
Then, “Give me freedom or give like the weasel.”
Then, “Give me freedom or give like a weasel.”
Then, “Give me freedom or think like a weasel.”
I skipped a few steps and I did not go all the way to the end, but I am sure you get the point. Eventually we could get all the way to Dawkins's "Methinks it is like a weasel" while maintaining function the entire time (although not necessarily beneficial function). Even Dawkins’s computer could do this type of phrase evolution… without an "ideal" phrase to compare with. Just program the computer to make random changes to single words within a phrase and then pick only those resulting phrases that actually make sense in English. The computer would quickly come up with all kinds of uniquely functional phrases. ( Back to Top )
Same Parts, Different Functions
But where would all these predefined words come from? Is there a parallel in nature to justify the use of preformed words in an evolutionary scenario? McDonald used preformed parts in the building of his mousetrap series. Can this be justified? Many have suggested that all the necessary parts exist within cells as parts of other systems of function. The genes for these parts are simply replicated for use in other evolving systems, sometimes with slight modifications. For example, Kenneth Miller, an evolutionary biologist from Brown University, points out that parts of a mousetrap could be used as a "tie clip, paper clip, toothpick, key chain or doorknocker." 7 Of course Miller is insinuating that this parallels real life where gene products that already do one job can be replicated and used in unique ways and combinations to do other jobs - even highly complex jobs. ( Back to Top )
It
is in fact true that the same or very similar genes can do very different jobs; not only within the same creature but in very different creatures as well.
This of course indicates a common evolutionary ancestor - or does it?
This hypothesis might seem like a possibility, but obviously this is not the only possibility, and maybe not even the most likely possibility at that. Is it too silly to suggest that the same author could have used the same words or codes in his writing of two different books or computer programs? Or, maybe two different authors could have used the same words, sentences, and even paragraphs to write their own books or computer programs?
So, when it comes to such similarities in the natural world, which scenario is most likely? As is usually the case, “The devil is in the details.” ( Back to Top )
Putting Parts Together in a Meaningful Way
Lets take another look at the phrase, "Methinks it is like a weasel." Lets assume that this is just one phrase in an entire book of phrases. Somewhere else in the book there is a phrase that reads, “Eating too much makes you fat.” Lets say that the word "much" happens to get duplicated. Before we continue further we must consider the odds of this happening.
It is true that duplication mutations do happen in real life, but they
are extremely rare events.
For mammals the average mutation rate is 2.2 base changes (character
changes) per year for a sequence that is one billion characters long.
Now, note that if a gene averages 10,000 bases (characters) in length,
this gives enough room for 100,000 genes in a genome a billion characters in
size. Of
course, most creatures do not have even close to 100,000 unique genes in their
genome. For
example, humans have only about 35,000 to 40,000 unique genes in a genome of
about three billion bases.4
The genes themselves occupy less than 10% of the total genomic
realestate.3,8
In fact, most mammals have a similar excess of non-coding DNA real estate
in their genomes.3
However, from the perspective of a single gene, it makes no difference
how many other genes there are in the genome. The average number of
generations needed to achieve even a single point mutation in a particular gene
would still be around 100,000 generations.5,6
With a generation time of 10 years, this is an average of one mutation
for a given gene every million years.
We can therefore assume that the average time needed to achieve a
duplication mutation for a given mammalian gene is also one million years at the
very least (since duplication mutations are much less common than single
character "point" mutations). Of
course, if there are a million individuals in a population, the "million
years" required to sustain one mutation in a particular gene in at least
one individual in that population would be realized in just one year.
With this background in mind, lets get back to our hypothetical phrase evolution. We have the phrase, "Methinks it is like a weasel" in our book - a book made up of many different phrases. If the word "much" happened to be duplicated from another phrase, "Eating too much makes you fat", then this word could be inserted into our "weasel phrase" in a functional way. Our newly mutated phrase could read, "Methinks it is much like a weasel." This mutation not only makes grammatical sense, but it also changes the meaning of the phrase just a little bit. However, what are the odds that this mutation would happen in the proper origin and insertion positions? If the mutation had placed the word "much" in another place it could have destroyed the function of the phrase altogether.
For example, the insertion could have caused the mutated phrase to read,
"Methinks it is like a much weasel."
While 30 million years or so is well within the proposed time frame for evolution, remember that this is a relatively short sequence. The real problems come when one remembers that each doubling of the minimum sequence length results in an exponential increase in the average time required to achieve success. In order to keep up with these odds, the population would also have to increase exponentially. The only problem with this is that population size is limited by environmental factors. Very quickly trillions upon trillions of years are required to evolve any novel function that has a minimum threshold requirement of more than one or two thousand amino acid residues working together at the same time in a fairly specific order relative to each other (see calculations). ( Back to Top )
All the Parts, But No Direction
So, just because the required parts needed to perform a particularly beneficial function in a cell already exist as parts of other functions, does not mean that these parts will just "self-assemble" to form a new beneficial function of high complexity whenever this function might be beneficial. The parts themselves simply do not know how to self-assemble beyond what information in contained preformed in the DNA. They do in fact have the potential to form many different collective interactions, but this is the problem. With the potential to interact in an almost infinite number of ways, no specific interaction is preferred over any other potential interaction. If left to themselves, the parts will not self-assemble to form very much of anything beyond a rather homogenous junk-yard collection of random parts and low-level functions.
It would be like putting all the parts needed for a watch into a bag and shaking the bag for a million years and expecting the parts to self-assemble into a working watch. It just doesn't happen without the outside informational input of a preformed code or an intelligent/deliberate action. Of course the powers of natural selection are supposed to solve this problem for evolutionists. But again, what happens to natural selection if the vast majority of possible arrangements of parts are equally non-beneficial from the perspective of a given system in a given environment?
At this point, the only thing left to sort through all the equally meaningless possibilities is a purely random change. Such a random walk simply takes more and more amounts of time to achieve success with each step up the ladder of minimum threshold requirements (i.e., "functional complexity") until, at relatively low levels of meaningful/functional complexity, the average random walk works its way into the trillions upon trillions of years of time. ( Back to Top )
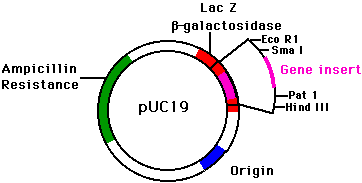 Now,
if these scenarios do not sound likely when it comes to real life, consider the
bacterial plasmid diagram depicted here. Notice the location of the gene
insertion (in the middle of the beta-galactosidase gene). How would a
researcher know if the desired gene was successfully inserted into the bacterial
plasmid?
Now,
if these scenarios do not sound likely when it comes to real life, consider the
bacterial plasmid diagram depicted here. Notice the location of the gene
insertion (in the middle of the beta-galactosidase gene). How would a
researcher know if the desired gene was successfully inserted into the bacterial
plasmid?
Interestingly enough, one of the ways to know that the insertion of a particular gene was successful is that this gene product is produced while at the same time some other pre-existing gene product, such as the pre-existing functional galactosidase enzyme, is not produced. The insertion of a gene in the middle of the galactosidase gene destroyed the function of the galactosidase gene.
Obviously then, random gene duplication events and
insertion events are rarely beneficial - especially when it comes to biosystem
functions that require greater and greater minimum threshold requirements. The
gap between what is and what might be beneficial at higher and higher levels of
functional complexity simply becomes to great to cross this side of a practical
eternity of time. (
Back to Top )
Also,
once a gene is duplicated, there is no reason why the entire gene would be
copied - functionally intact.
The duplication might only include a portion of the gene and this portion
might not make sense.
For example, instead of our previous duplication mutation reading
"much" it could have read, "o muc" or "akes you fa."
What then are the odds that any one duplication event will duplicate
something functionally intact?
It seems like the odds are pretty slim at best.
Then, even after something functional is duplicated, what are the odds
that this functional duplication will be inserted in a functional and beneficial
way in another location?
Again, as previously discussed, the odds are pretty slim and they get
more and more slim, in an exponential manner, with increasing minimum threshold
requirements.
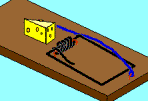
Now, take another look at McDonald’s mousetrap example. If we have a mousetrap and we need some cheese to make it better, what are the odds that cheese will come our way? What if a duplication mutation brought along some "heese" and placed it in just the right place? Close, but close is not good enough here. “Heese” has no meaning at all, much less the "beneficial" meaning of "cheese." Now, we are stuck waiting for another ten millions years for some real c-heese to come along. Of course the odds are that with each mutation that affects our evolving mousetrap, the cheese code (or more likely - a partial cheese code) will come along once every ten million years or so. The problems are not yet over of course. The functional cheese could mutate into the wrong spot in our mousetrap gene. If our mousetrap gene is 1,000 characters long, and the cheese will only work in 5 different positions, that means that only once in 200 cheese code arrivals will the placement actually be beneficial. This means that it would take billions of years for the cheese code to evolve effectively into our mousetrap gene - on average (even in a fairly large population).
Now, what happens when the "gap" is wider than "cheese"? - a gap where a specific phrase of 20 or 30 characters is required? Again, with each linear increase in the minimum gap size, the random walk/selection time increases exponentially. ( Back to Top )
McDonald has tried to show that Behe’s mousetrap is not "irreducible", but it turns out that each of the steps proposed by McDonald might be individually complex to such an irreducible degree that evolution stalls out at the very first step. The mindless processes of evolution simply cannot overcome the average time of random walk required to realize even one, much less all of McDonald's proposed steps in a reasonable amount of time. And, this problem gets exponentially worse with greater and greater minimum threshold requirements. ( Back to Top )
As with other paper theories, like McDonald's, no one has ever succeeded in getting such scenarios off the paper and into real life demonstrations. There are many examples of very simple functions evolving where the function realized is dependent upon the interference of a pre-established function or interaction (i.e., antibiotic resistance and the like). Moving up the ladder of functional complexity there are far fewer examples of single protein enzymes evolving, but there are actually a few examples, like lactase and nylonase etc., evolving at this level. However, when one moves up the ladder of complexity to functions that require more than a few hundred (say one or two thousand) fairly specified amino acids all working together at the same time (i.e., bacterial motility systems like the flagellum, cilium, undulating membranes, transcription, translation, pinocytosis, vision systems, etc.), there simply are no real time examples - none. There aren't even examples of evolution across even one step of the proposed evolutionary pathways for such systems. It just doesn't happen.
Behe accurately points this problem out and makes the problem intuitively obvious with his mousetrap illustration as well as many other such illustrations. McDonald has tried to muddy the waters a bit, but even on paper his position has serious statistical problems. Then, when it comes to real life predictive value / demonstration, there simply is no support for McDonald's "just-so" assertions when it comes to explaining how many types of biosystems could ever have evolved. ( Back to Top )
Dawkins, Richard. The Blind
Watchmaker, 1987.
Behe, Michael J. Darwin’s Black
Box, The Free Press, 1996.
Lewin, Benjamin. Genes
V, Oxford University Press, 1994.
Lemonick, M. Gene Mapper,
Time, Vol. 156, No. 26, pp110, 2001.
Dugaiczyk,
Achillies. Lecture
Notes, Biochemistry 110-A, University California Riverside, Fall 1999.
Ayala, Francisco J. Teleological
Explanations in Evolutionary Biology, Philosophy of Science, March, 1970, p.
3.
Miller, Kenneth R., Finding
Darwin’s God, HarperCollins Publishers, 1999.
F.
Flam, Hints of a language in junk DNA, Science 266:1320, 1994.
http://udel.edu/~mcdonald/mousetrap.html, June, 2002.
Hayashi Y, Sakata H, Makino Y, Urabe I, Yomo T., Can an arbitrary sequence evolve towards acquiring a biological function? J Mol Evol. 2003 Feb; 56(2):162-8 ( Link )
. Home Page . Truth, the Scientific Method, and Evolution
.
. Maquiziliducks - The Language of Evolution . Defining Evolution
.
.
Evolving
the Irreducible
.
.
.
.
.
. DNA Mutation Rates . Donkeys, Horses, Mules and Evolution
.
.
. Amino Acid Racemization Dating . The Steppingstone Problem
.
.
Since June 1, 2002